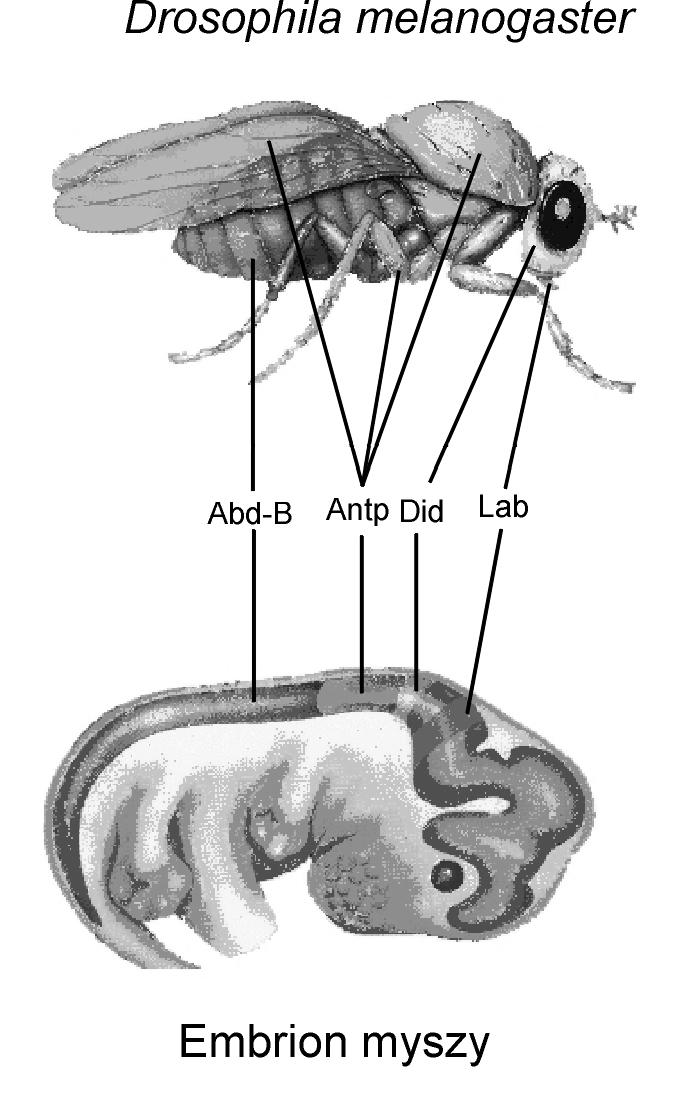
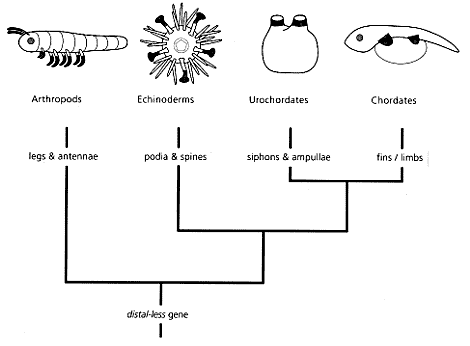
'Wspinaczka na szczyt nieprawdopodobieństwa', Richard Dawkins napisał:

„
(....)Richard DawkinsMogłoby się wydawać, że twierdzeniu temu przeczą intrygujące wyniki eksperymentów, jakie przeprowadzili ostatnio w Szwajcarii naukowcy pracujący pod kierownictwem Waltera Gehringa. Przedstawię je pokrótce i udowodnię, że w istocie wcale nie są sprzeczne z naszą konkluzją. Zanim rozpocznę, muszę przeprosić czytelników za zwariowaną konwencję nazywania genów przez genetyków. Gen nazwany eyeless (po polsku - bezoki) u muszki owocowej Drosophila w istocie odpowiedzialny jest za powstanie oczu (prześliczne, prawda?!)! Powód tego figlarnie mylącego przykładu terminologicznej sprzeczności jest w rzeczywistości Richard Dawkinsdość prosty, a nawet interesujący. Otóż funkcję, jaką pełni określony gen, można rozpoznać wówczas, gdy dzieje się z nim coś złego. Kiedy gen, o którym mówimy, psuje się (czyli mutuje), u muchy nie rozwijają się oczy. Miejsce na chromosomie, w którym znajduje się ten gen, nazywane jest więc eyeless locus (locus to po łacinie miejsce, genetycy określają w ten sposób miejsce na chromosomie, w którym zlokalizowane są różne formy jednego genu). Mówiąc jednak o genie eyeless, mamy zwykle na myśli jego normalną, nieuszkodzoną wersję znajdującą się w tym locus. Stąd paradoks, że gen „bezoki” decyduje o powstawaniu oczu. To tak, jakby głośnik nazwać „wyciszaczem”, kiedy odkryje się, że po jego odłączeniu radio cichnie. Już przestaję. Mam tylko ochotę nazwać ten gen „eyemaker (po polsku - „okorób”), ale to chyba też specjalnie nie ułatwiłoby sprawy. Nie mogę jednak nazywać go eyeless, posłużę się więc skrótem ey.
Jak powszechnie wiadomo, choć wszystkie komórki każdego organizmu zawierają wszystkie geny, tylko niewielka ich część jest wykorzystywana (ulega ekspresji) w poszczególnych częściach ciała. Dlatego właśnie wątroba różni się od nerek, chociaż ich komórki zawierają dokładnie takie same geny. U dorosłej muszki owocowej gen ey zwykle ulega ekspresji tylko w komórkach głowy - dlatego oczy rozwijają się właśnie tam. George Haider, Patrick Callaerts i Walter Gehring opracowali procedurę powodującą powstawanie oczu także w innych rejonach ciała tego owada. Hodując larwy muszek w specjalny sposób, doprowadzili do tego, że gen ey uległ ekspresji w miejscach, w których normalnie powstają czułki, skrzydła i odnóża. Co jeszcze zabawniejsze, uzyskali dorosłe muchy z całkowicie rozwiniętymi oczami złożonymi na skrzydłach, odnóżach, czułkach i w innych miejscach (ryc. 5.29). Choć nieco mniejsze, oczy te mają całkiem normalną budowę i składają się z wielu właściwie ukształtowanych ommatidiów. A nawet działają. Nie wiemy, co prawda, czy muchy rzeczywiście coś przez nie widzą, ale rejestracja aktywności elektrycznej nerwów leżących u podstawy ommatidiów dowodzi, że są one wrażliwe na światło.
To jest godny uwagi fakt numer jeden. Fakt numer dwa jest jeszcze bardziej interesujący. Znany jest gen mysi, zwany small eye (po polsku - drobnooki) i gen ludzki, zwany aniridia (po polsku - bez tęczówki). Ich nazwy są także odzwierciedleniem negatywnej konwencji przyjętej przez genetyków: mutacyjne uszkodzenie tych genów powoduje zmniejszenie bądź brak oczu lub ich części. Rebecca Quiring i Uwe Waldorf z tego samego szwajcarskiego laboratorium stwierdzili, że oba te geny ssaków są niemal identyczne pod względem sekwencji DNA z genem ey muszki owocowej. Oznacza to, że ten sam gen przeszedł cały łańcuch przodków aż do obecnie żyjących zwierząt tak daleko ze sobą spokrewnionych jak owady i ssaki. Co więcej, u obu tych bardzo odległych grup zwierząt gen ten ma decydujące znaczenie w rozwoju oczu. Fakt numer trzy jest wręcz zdumiewający. Haiderowi, Callaertsowi i Gehringowi udało się wprowadzić gen mysi do zarodków muszek owocowych. Ku wielkiemu zdumieniu okazało się, że spowodował powstanie ektopowych oczu u Drosophila. Rycina 5.29 przedstawia niewielkie oczy złożone, jakie rozwinęły się na odnóżu muszki w wyniku indukcji mysim odpowiednikiem genu ey. Warto przy tym zwrócić uwagę, że powstało wówczas owadzie oko złożone, a nie oko charakterystyczne dla myszy. Mysi gen po prostu uruchomił mechanizmy embriogenetyczne odpowiedzialne za powstawanie oczu u muszki owocowej. Geny o niemal takiej samej sekwencji DNA jak gen ey znaleziono także u mięczaków, morskich robaków zwanych wstężnicami i u strzykw. Być może okaże się, że ey występuje u wszystkich zwierząt i podanie pewnej jego wersji pochodzącej z jednej grupy zwierząt przedstawicielowi jakiejś innej, bardzo odległej grupy spowoduje u niego rozwój oczu. Richard Dawkinsinduktorem oczu widocznych na najniższym zdjęciu był gen mysi.
Co wyniki tych spektakularnych doświadczeń znaczą dla konkluzji tego rozdziału? Czy myliliśmy się mówiąc, że oczy wykształciły się czterdzieści razy niezależnie od siebie? Nie sądzę. W mocy pozostaje przynajmniej duch twierdzenia, że oczy ewoluują łatwo i szybko. Wyniki te świadczą natomiast prawdopodobnie o tym, że wspólny przodek muszek owocowych, myszy, ludzi, strzykw i innych stworzeń miał oczy. Ten odległy przodek dysponował zapewne jakimś rodzajem wzroku, a o powstaniu jego oczu, niezależnie od ich postaci, prawdopodobnie zadecydowała sekwencja DNA podobna do tej, jaką ma dzisiejszy gen ey. Współczesne postaci różnego rodzaju oczu, szczegóły budowy ich siatkówek, soczewek czy zwierciadeł, wybór między okiem prostym a złożonym oraz między okiem apozycyjnym a różnymi odmianami oka superpozycyjnego są jednak rezultatem niezależnej i szybkiej ewolucji. Możemy się o tym przekonać, obserwując przypadkowe - można wręcz powiedzieć: kapryśne - rozrzucenie najróżniejszych rozwiązań, pomysłów i systemów w całym królestwie zwierząt. Mówiąc krótko: często badane zwierzę ma oczy przypominające dużo bardziej oczy swego bardzo odległego krewnego niż bliskiego kuzyna. Konkluzji tej w niczym nie narusza konstatacja, że wspólny przodek wszystkich zwierząt najprawdopodobniej miał jakieś oczy, i że rozwój zarodkowy oczu wszystkich zwierząt ma ze sobą wystarczająco dużo wspólnego, by można go było zaindukować tą samą sekwencją DNA. (….)”
A więc jak widzisz gen PAX-6 stoi u samej podstawy rozwoju oczu u wszystkich obdarzonych oczami organizmów na ziemi. Teraz się zastanów, w jaki sposób duża ilość innych „HOMOLOGICZNYCH” genów zaczęło STEROWAĆ rozwojem NIEhomologicznych oczu? Pomyśl: wszystkie kręgowce mają specyficzną cechę, mają odwróconą siatkówkę w oku:
http://bioslawek.wordpress.com/2012/01/ ... kregowcow/

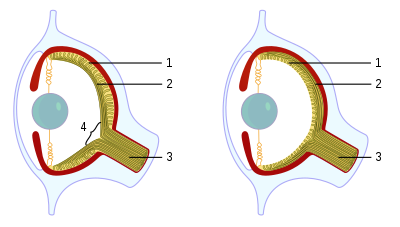 1)Kręgowiec, 2) Osmiornica
1)Kręgowiec, 2) Osmiornica
Choć oczy głowonogów są tak podobne do oczu kręgowców, to jednak mają „normalną” siatkówkę. NIE odwróconą, jak kręgowce. Jaki z tego wniosek dla ewolucjonisty? Kręgowce musiały mieć WSPÓLNEGO PRZODKA, u którego wyewoluowała odwrócona siatkówka, więc oko kręgowców ewoluowało osobno niż oko głowonogów. Obydwa rodzaje oczu, myśląc ewolucyjnie, musiały wyewoluować NIEZALEŻNIE od siebie, STARTUJĄC od komórek światłoczułych, gdzie już działał gen PAKS-6! A więc skoro oko kręgowca i głowonoga wyewoluowało, zdaniem ewolucjonistów, w wyniku ewolucji konwergentnej (zbieżnej), to oczy kręgowców i głowonogów nie mogą stanowić struktur homologicznych. Czyli pochodzących od WSPÓLNEGO PRZODKA, który żył już po nabyciu genu PAX-6.

Czego w takim układzie powinni spodziewać się genetycy badający ewolucję oczu? Ano tego, że geny sterujące rozwojem oczów kręgowców i głowonogów, które, patrząc ewolucyjnie, musiały wyewoluować już po powstaniu genu P-AX-6 powinny się od siebie RÓŻNIĆ, powinny nie być HOMOLOGICZNIE, a one w kilkudziesięciu procentach SĄ „HOMOLOGICZNE” (takie same, lub bardzo podobne)! Czy uważasz naprawdę, że ewolucjoniści w tym przypadku nie mają się z czego tłumaczyć?
Należy tutaj jeszcze raz podkreślić, że oczy kręgowców i głowonogów, to nie jedyne, których rozwój dostarcza ewolucjonistom tylu nierozwiązanych problemów, ponieważ tak się dzieje w przypadku WSZYSTKICH zbadanych organizmów obdarzonymi oczami, których rozwój startuje od genu PAX-6. A badania trwają dalej. No i jak się mogłeś przekonać identyczne problemy nie tylko dotyczą oczu, ale też kończyn, czy innych części ciała u różnych stworzeń. A nauka prze do przodku i bada coraz więcej organizmów pod tym kątem, choć takie badania są mozolne i niewdzięczne. Faktem pozostaje jednak prawda, że teoria ewolucji nie zyskuje na rozkwicie nauk zajmujących się biologią rozwoju i genetyką rozwoju.
Na koniec od razu zdementuje jeszcze inną dezinformację szerzoną przez ewolucjonistów. Żeby jakoś pominąć problemy związane z „HOMOLOGICZNYMI” genami kodującymi NIE-HOMOLOGICZNE ORGANY zwracają oni uwagę na podobieństwa opsyn u różnych organizmów. Opsyny to grupa białek, które na poziomie biochemicznym powodują, że się widzi.
http://pl.wikipedia.org/wiki/Opsyny „Opsyny to grupa światłoczułych sprzężonych z białkami G receptorów błonowych o masie cząsteczkowej 35-55 kDa, należących do rodziny białek retinylydynowych. Opsyny występują w warstwie receptorowej siatkówki. Pięć klasycznych grup opsyn jest zaangażowanych w proces widzenia, pośrednicząc w przekształceniu fotonów światła w sygnał elektrochemiczny, pierwszym etapie transdukcji sygnału wzrokowego. Inne białko z grupy opsyn, melanopsyna, zaangażowane jest w rytmy dobowe i odruch źreniczy, ale nie bezpośrednio w proces widzenia.”. Twierdzi się też, że biochemiczne procesy związane z widzeniem, które zachodzą w komórkach światłoczułych, wyewoluowały kilka razy niezależnie, choć są to uklady NIEREDUKOWALNIE ZŁOZONE.
Pomińmy na razie tą nieredukowalną złożoność procesów biochemicznych odpowiedzialnych za widzenie i powróćmy do genów sterujących rozwojem oka. Jak napisał Dawkins, gdy zabraknie genu PAKS-6 oko się nie wykształci. Tak naprawdę rozwój oka nie następuje, lub wykształca się oko ślepe, gdy zabraknie jakiegokolwiek genu w KASKADZIE, jaka zachodzi w SIECI GENETYCZNEJ zawiadującej rozwojem oka. Więc jak takie sieci mogły ewoluować niezależnie od siebie tyle razy? To już kolejny problem ewolucjonistów.
Ale powróćmy do problemu kodowania NIE-HOMOLOGICZNYCH struktur przez „HO-MOLOGICZNE” geny. Nawet jeśli się założy, że procesy biochemiczne odpowiedzialne za widzenie, w których biorą udział opsyny, wyewoluowały już na etapie komórek światłoczułych, jeszcze przed pojawieniem się genu PAX-6, to przecież taka możliwość (zaraz pokażę dlaczego było to niemożliwe) nie wyjaśnia, jak „HO-MOLOGICZNE” geny zaczęły już po pojawieniu się tych procesów biochemicznych, podobnych u wszystkich widzących organizmów oraz genu PAX-6 sterować rozwojem NIE-HOMOLOGICZNYCH struktur (oczu).
Na koniec powrócę do procesów biochemicznych odpowiedzialnych za widzenie, które zachodzą w komórkach światłoczułych. Z punktu widzenia biochemii proces widzenia wygląda dość skomplikowanie.
Gdy światło dociera do siatkówki, foton reaguje z cząsteczką (białkiem) zwaną 11-cis-retinalem, która w ciągu kilku pikosekund przemienia się w trans-retinal. Zmiana kształtu (konformacji) cząsteczki retinalu wymusza zmianę kształtu białka- rodopsyny, z którym retinal jest ściśle związany. Z metamorfozą białka idzie w parze zmiana jego funkcji. Białko nazywane teraz metarodopsyną II przyczepia się do innego białka zwanego transducyną. Przed przylgnięciem do metarodopsyny II , transducyna mocno wiązała małą molekułę zwaną GDP. Lecz gdy transducyna reaguje z metarodopsyną II , GDP zostaje uwolniony, a z transducyną wiąże się molekuła GTP (GTP jest blisko spokrewnione z GDP, ale różnią się one istotnie).
W tym momencie GTP-transducyno-metarodopsyna II wiąże się z białkiem zwanym fosfodiestrazą, które znajduje się w wewnętrznej błonie komórki. Po związaniu z metarodopsyną II i jej otoczeniem, fosfodiestraza nabywa zdolności „odcinania” cząsteczki zwanej cGMP. Początkowo w komórce znajduje się wiele cząsteczek cGMP, ale fosfodiestraza obniża ich koncentrację tak, jak wyjęcie korka powoduje obniżenie poziomu wody w wannie.
Inne białko błonowe, wiążące się z cGMP zwane jest kanałem jonowym. Działa on jak brama wjazdowa, która reaguje liczbę jonów sodu w komórce. Zwykle kanał jonowy pozwala na wpływanie jonów sodu do komórki, gdyż osobne białka aktywnie wypompowują je z powrotem. Wspólne działanie kanału jonowego i pompy utrzymuje w komórce w miarę stały poziom jonów sodu. Gdy ilość cGMP w komórce zostaje zredukowana z powodu rozczepienia przez fosfodiestrazę, kanał jonowy zamyka się powodując obniżenie dodatnio naładowanych jonów sodu w komórce.
To z kolei powoduje zachwianie potencjału elektrycznego w komórce i wysyłanie impulsu do mózgu przez nerw wzrokowy. Efektem tego, po zinterpretowaniu przez mózg, jest widzenie.
Poniżej ilustracje obrazujące powyżej opisany proces.





Gdyby opisane wyżej reakcje były jedynymi zachodzącymi w komórce, to zasoby 11-cis-retinalu, cGMP i jonów sodu szybko by się wyczerpały. Coś (jakieś procesy) musi wyłączać białka, które były włączone i przywracać komórce jej stan pierwotny. Uczestniczy w tym kilka mechanizmów biochemicznych. Po pierwsze: w ciemnościach kanał jonowy oprócz jonów sodu wpuszcza jeszcze do komórki także jony wapnia. Są one wypompowywane z powrotem przez inne białko tak, żeby utrzymywała się stała koncentracja wapnia. Gdy opada poziom cGMP , za przyczyną zamknięcia kanału jonowego, obniża się również koncentracja jonów wapnia. Enzym fosfodiestraza, który niszczy GMP powodując spadek jego stężenia w komórce, spowalnia swoje działanie przy mniejszej koncentracji wapnia
Po drugie: gdy zaczyna się obniżać poziom wapnia, białko zwane cyklazą guanylową rozpoczyna ponowną syntezę cGMP.
Po trzecie: w trakcie tego procesu metarodopsyna II zostaje zmodyfikowana przez enzym zwany kinazą rodopsynową . Zmodyfikowana rodopsyna wiąże się wówczas z białkiem zwanym jako arestyna, które nie dopuszcza do aktywacji przez rodopsynę większej ilości transducyny
W komórce więc zachodzą reakcje zapobiegające wzmocnieniu sygnału wywołanego przez pojedynczy foton.
Trnas-retinal zostaje ostatecznie uwolniony z rodopsyny i musi ponownie być przemieniony w w 11-cis-retinal, następnie zostaje związany przez rodopsynę, wracając do punktu wyjściowego kolejnego cyklu procesu widzenia .
Aby to osiągnąć trans-retinal jest najpierw chemicznie przekształcany przez enzym w trans-retinol -formę posiadającą dwa dodatkowe atomy wodoru.
Drugi enzym następnie przekształca tą molekułę w 11-c-s-retinol.
W końcu trzeci enzym usuwa wcześniej dodane dwa atomy wodoru, tworząc 11-sic-retinal. Cykl się zamyka.
Teraz zastanów się czy taki nieredukowalnie złożony i skomplikowany proces mógł powstać STOPNIOWO na drodze ewolucji?
Podsumujmy naszą dyskusję. Ewolucjoniści w przypadku ewolucji oczu (bo tylko ten przypadek rozpatrywałem) borykają się z następującymi nieprzebytymi problemami:
1) Nie potrafią wyjaśnić, jak „HO-MOLOGICZNE” geny mogą sterować rozwojem NIE-HOMOLOGICZNYCH organów.
2)W jaki sposób wyewoluowały wiele razy niezależnie od siebie złożone sieci sterujące rozwojem kilkudziesięciu typów oczu.
3) W jaki sposób mógł wyewoluować nieredukowalnie złożony proces widzenia na poziomie biochemicznym.